![]()
Диметродон (лат. Dimetrodon) — род пермских пеликозавров (синапсидов), живших 280—265 млн лет назад на территории современных США и Германии.
История изучения[1]
Раннее изучение
Первоначально останки диметродона описал Эдвард Дринкер Коуп в 1870-ых. Он получил их вместе с коллекцией других пермских тетраподов, полученных из формации Красные грады в Техасе. Передали их Коупу коллекционер Якобс Болл, геолог У. Ф. Камминс и палеонтолог Чарльз Стернберг Большая часть этих образцов сейчас находится в Американском музее Естественной истории или в музее Уокера Чикакгского университета.
Стернберг отправил несколько своих образцов немецкому палеонтологу Фердинанду Бройли из Мюнхенского университета, однако он не так хорошо изучил их, как Коуп. Соперник Эдаврда, Чарльз Марш, также собрал несколько костей диметродона, однако он отдал их в музей Уокера.
Впервые наименование Dimetrodon использовал Коуп в 1878 году, выделив три вида — D. incisivus, D. rectiformis и D. gigas.


Одна из самых известных картин с диметродоном
Однако первое описание останков диметродона было в 1875 году, когда Коуп описал клепсидропса C. limbatus. Это животное было обнаружено в той же местности, что и диметродон, и в конце XIX-начале ХХ века большинство останков пеликозавров приписывали диметродону или клепсидопсу. В 1940 году вышла публикация, в которой говорилось, что C. limbatus является на самом деле видом диметродона.
Первым описанным синапсидом с парусом был клепсидропс C. natalis, также выделенный Коупом. Он посчитал парус плавником и сравнивал с гребнем ящерицы василиска. Паруса D. incisivus и D. gigas не сохранились, однако у образца D. rectiformi сохранились удлинённые невральные шипы. Однако Коуп прокомментировал в 1886 году, что назначение паруса сложно представить. По его словам, если животное не вело водный образ жизни, то парус или плавник мешал бы движениям, а конечности и когти не были достаточно большими, чтобы вести древесный образ жизни, как у василиска.
ХХ век
| Много меньше прожили загадочные пеликозавры, по современным представлениям, близкие к предкам динозавров. Появились они в карбоне и на протяжении перми господствовали на территории Северной Америки, были там всем сразу - и хищниками, и травоядными, как в то время тероморфы в Восточном Европе. Самые странные имели 'парус' из перепонки, натянутой вдоль спины на длинных, иногда шипастых остистых отростках, как у диметродона, или 'двумера', на реконструкции. Полагают, что гребень мог служить примитивным терморегулятором, по-разному располагаясь относительно ветра и солнца. Правда, трудно придумать более несуразный регулятор. |
В начале ХХ века Э. Кейс провёл серьёзное исследование диметродона, выделив несколько новых видов. Ему помог с финансированием Институт Карнеги, предоставив также материал из различных музеев Америки. Многие образцы были описаны Копом, прославившимся тем, что описывал целые рода лишь по фрагментам, однако он не обратил на эти останки много внимания.
Начиная с концов 1920-ых Альфред Ромер провёл повторное изучение многих образцов демитродона, выделив ещё несколько видов. В 1940 году Ромер и Ллевеллин Прайс провели «Обзор пеликозавров», в котором просмотрели большинство синапсид, описанных Копом. Большинство результатов этого исследования актуальны до сих пор.


Старая реконструкция вида Dimetrodon incisivus
После публикации Ромера и Прайса последовало открытие многочисленных образцов диметродона вне Оклахомы и Техаса. Так, в 1966 году были обнаружены небольшие фрагменты на территории Юты, а в 1969 году останки нашли в Аризоне. В 1975 году Олсон сообщил о находке диметродона в Огайо. В 1977 Берман описал на основе материала из Нью-Мексико вид D. occidentalis («западный»), к которому также отнесли останки из Юты и Аризоны.
До этих открытий считалось, что Техас и Оклахому от остальных территорий отделял Серединноконтинентальный морской путь, благодаря чему в западной Северной Америке обитал меньший сфенакодон. Новые находки хоть и не опровергают наличие внутреннего моря, но указывают на его ограниченность и то, что оно не было преградой для расселения диметродона.
Новейшие исследования
В 2001 году в нижней перми Бромакера, Тюрингия, был обнаружен новый вид диметродона, названный D. teutonis. Некоторые исследователи говорят, что диметродон обитал не только в Германии, но и на территории Южной Америки и России.
Описание
| Даже молодой диметродон, существо совсем не микроскопических размеров, наелся в тот раз до отвала, и после его трапезы на костях оставалось ещё немало съестного… но, видимо, не настолько много, чтобы кормить всех местных плотоядных в течение нескольких дней, поскольку, когда хищник всё же добрался до знакомого места, то не увидел ничего кроме вытоптанной земли, нескольких клочков шкуры и пары слишком крупных костей, что не смогли уместиться ни в чьём объёмистом желудке. |


Dimetrodon macrospondylus
У диметродона была довольно большая голова с крепкими челюстями, усеянными острыми зубами. Вероятно, он был активным охотником: охотился на амфибий, пресмыкающихся и рыбу. Своими передними зубами диметродон держал жертву и разрывал ее на части. Задние зубы были загнуты назад, с их помощью животное разрывало на части мелких существ и пережевывало большие куски мяса. Туловище диметродона имело бочкообразную форму. Наиболее яркой чертой диметродона является парус из натянутой на выросты спинных позвонков кожи. Подобные паруса развились у различных доисторических животных (амфибии платигистрикса, пеликозавров эдафозавра и секодонтозавра, динозавра спинозавра) и служили терморегуляторами. По другим версиям парус использовался в брачных играх, служил камуфляжем среди вертикальных стеблей растений или настоящим парусом при плавании. «Парус» развивался постепенно в течение жизни, известны находки молодых особей диметродона с невысокими остистыми отростками. Молодые особи, по-видимому, обитали в зарослях по берегам водоемов, как и взрослые. Тем не менее, бытующее в популярных книгах изображение диметродона на фоне пустыни представляется нереальным — не являясь рептилией он не мог бы выжить в жаркой сухой местности.
Череп[2]
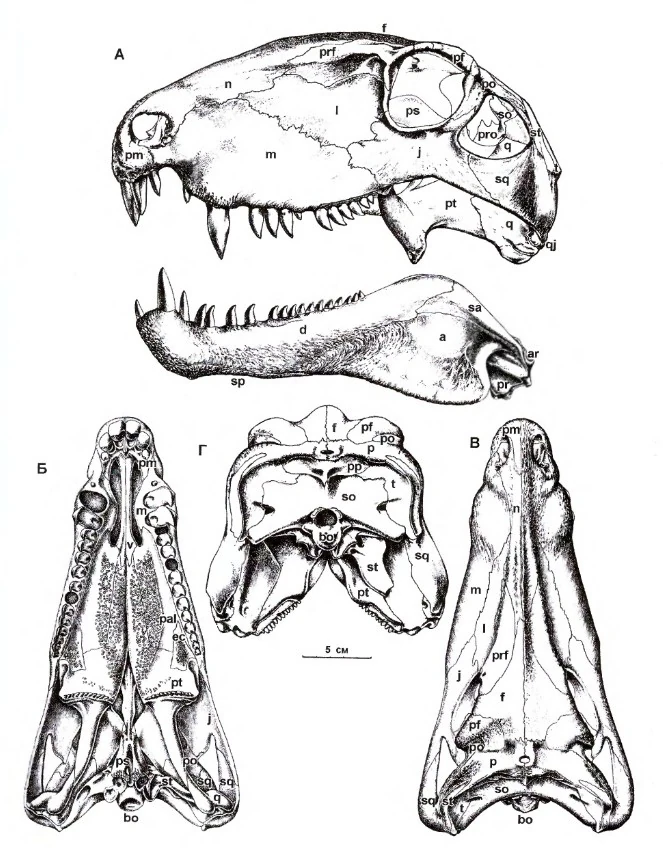

Череп диметродона высокий, сжатый с боков. Премаксилла отделена от челюсти глубокой диастемой. Передний край сильно выпуклый. Небольшие ноздри находятся в передней части морды. Слёзная кость не доходит до ноздрей. Глазницы находятся в задней части черепа. На премаксилле находится три зуба, второй зуб верхней челюсти имеет клыковидную форму, увеличен, изогнут, с зазубренными краями. Зубцы очень тонкие, как у альбертозавра. У их оснований находились небольшие пустоты, снижающие нагрузку на зубы, но не спасающая от повреждений зубцов. У D. teutonis не было зазубрин, однако края были такие же острые. Нижний клык входит в диастему между премаксиллой и челюстью. Зубы обеих челюстей перед клыками редуцированны, за ними — постепенно уменьшаются в размерах. По форме зубы диметродона и его родственников напоминают капли, что помогает отличить сфенакодонтов от остальных ранних синапсид.
Исследования 2014 года показали, что диметродоны проводили своего рода гонку вооружений. Мелкие D. milleri не имели зазубрин на зубах, так как охотились на мелкую добычу. По мере увеличения видового разнообразия и размеров, зубцы изменяли свою форму. У D. limbatus зубцы имели режущую форму, как у секодонтозавра. У крупного D. grandis зубцы имели такую же форму, как у акул и тероподов. Таким образом, диметродоны не только увеличивали размеры по мере эволюции, но и изменяли привычные приспособления для охоты.


От завропсид диметродон отличается наличием инфратемпоральной фенестры. У рептилий было две фенестры, либо они отсутствовали вообще, в то время как у синапсид было всего одно такое отверстие. У диметродона были необычные признаки перехода от ранних тетропод к млекопитающим, как гребни на задней части нижней челюсти и внутри носовой полости.
На внутренней части носовой полости находились специальные гребни, насотурбины. Они могли поддерживать хрящи, увеличивающие площадь обонятельного эпителия. Эти гребни меньше, чем у млекопитающих и более поздних синапсид, у которых насотурбины являются возможным признаком теплокровности. На них могла находиться слизистая оболочка, нагревающая и увлажняющая поступающий воздух. Таким образом, диметродон был частично теплокровным животным.
Ещё одной особенностью диметродона является выступ в задней части челюсти, называемый отражённой пластинкой. Она находится на суставной кости, соединённой с квадратной костью, вместе образующими челюстной сустав. У более поздних синапсид отростки суставной и квадратной кости отсоединились от челюстного сустава, образовав кость среднего уха — молоточек. отражённая пластинка позднее развилась в барабанное кольцо, поддерживающее барабанную перепонку у современных млекопитающих.
Хвост
Долгое время диметродона представляли как животное с коротким хвостом, так как всего было известно 11 ближайших к телу хвостовых позвонков, которые сильно сужались по мере отдаления от таза, а у первых описанных останков хвост вообще отсутствовал, либо был не в лучшем состоянии. Только в 1927 году был открыт полный хвост диметродона, состоящий из 50 позвонков. Он составлял большую часть длины тела и служил балансиром при передвижении.
Парус
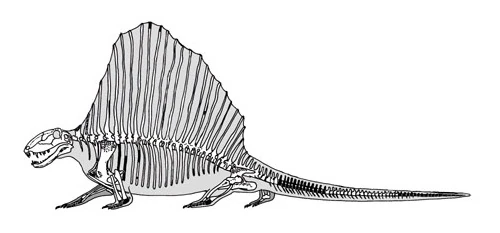
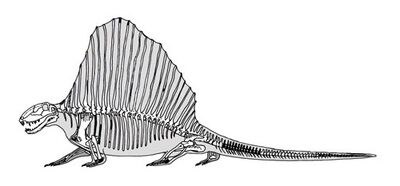
Скелет D. loomisi
Одна из самых ярких и выдающихся черт диметродона — высокие остистые отростки спинных и шейных позвонков. Со времен открытия рода их изображали по-разному: как просто торчащие шипы, так и полностью обтянутый кожей «парус», или даже горб. Парус достигал в высоту до одного метра. Верхушки невральных шипов были покрыты роговыми чехликами. Каждый невральный шип имеет уникальную форму, такая дифференциация даже получила название «диметродонтовая». Возле тела позвонка шипы имеют прямоугольную форму, по мере удаления от него принимая форму восьмёрки. Считается, что такая форма укрепляла отростки и мешала переломам. Известна одна особь D. giganhomogenes с полностью прямоугольными невральными шипами, однако возле центра всё ещё присутствуют следы «восьмёрок». Скорее всего, эти изменения связаны с возрастом особи. Микроанатомия каждого шипа позволяет разглядеть место прикрепления к мышцам и место перехода в парус. Нижняя, проксимальная часть шипа имеет шероховатую поверхность. Возможно, к ней крепились эпаксальные и гипаксальные мышцы, а также сеть соединительной ткани, так называемые волокна шарпи. Дистальная часть шипов гладкая, однако надкостница пронизана многочисленными бороздками, возможно, при жизни в них располагались кровеносные сосуды. Многослойная пластинчатая кость, составляющая большую часть поперечного сечения невральных шипов, содержит много линий роста, по которым можно определить возраст каждой особи на момент смерти.
Через все позвонки проходит необычный желобок. Раньше считалось, что в нём располагались кровеносные сосуды, но так как их следы отсутствуют внутри костей, возможно, что желобок предназначался для чего-то другого, а количество сосудов в парусе было гораздо меньше, чем считалось.
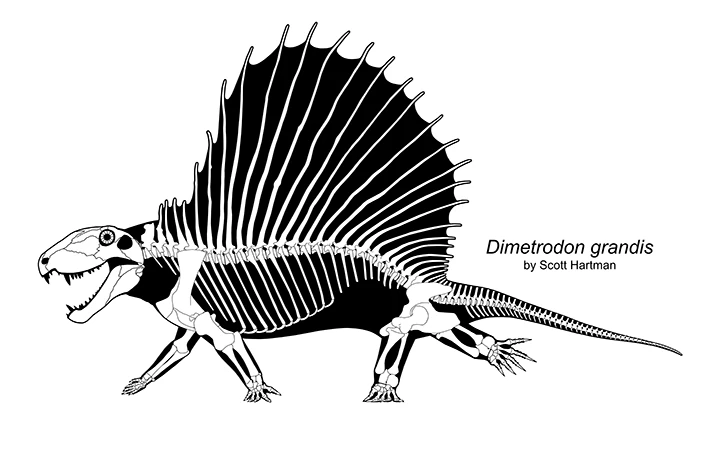
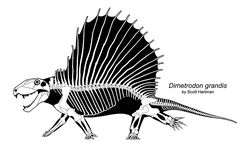
Современная реконструкция скелета диметродона. Автор — Скотт Хартман
Изучение патологий показало, что некоторые невральныеные шипы сломались, а затем исцелились. Это наводит на мысль, что по крайней мере часть отростков была обтянута парусом (или подобной тканью), которая удерживала их на месте после повреждения, в следствии чего они могли заживать. Но они также заметили, что вершины шипов часто согнуты, иногда очень сильно, что предполагает, что вершина шипов не входила в парус. Дальнейшее подтверждение этой конфигурации «выход из паруса» происходит из поверхностной текстуры отростков. Они были шероховатыми в том месте, где крепились к мышцам спины, далее становясь все более гладкими, как раз там, где они, скорее всего, становились шипами, торчащими из паруса.[3] Выросшая на месте переломов кортикальная кость сильно васкуляризирована, что говорит о наличие определённого числа мягких тканей и сосудов на парусе.
Остальной посткраниальный скелет
Плечевая кость очень широкая в нижней половине, с энтэпикондилярным отверстием. Сочленовные поверхности костей хорошо развиты.
Кожа
Отпечатки кожи диметродона неизвестны, поэтому говорить о какой-то определённой текстуре сложно. У более поздних синапсидов, вроде эстемменозуха, кожа была гладкой, с большим количеством желёз. Однако у более примитивных ранних варанопсеид, таких как аскендонан, удалось обнаружить отпечатки чешуи. Не исключено, что диметродон имел чешуйчатую текстуру на нижней стороне тела, а с боков и сверху его кожа была гладкой, как у терапсид.
Походка
| Он был размером с мою ладонь и выглядел почти как человеческая рука, но если бы большой палец заменили другим. |


Диметродон традиционно изображается с развалистой «ящеричной» походкой с волочащимся по земле брюхом, однако недавно были обнаружены дорожки, принадлежащие диметродону или близкому к нему синапсиду, показывающие животное, передвигающееся с более выпрямленными ногами, держа живот и хвост полностью свободными от земли.
Конечно, диметродон мог разлечься, когда захочет. Однако во время ходьбы и бега его конечности всё-таки имели полувыпрямленное положение, из-за чего диметродон мог быть быстрее своих жертв (амфибий и более мелких синапсид).
Виды и ареал


Скелет Dimetrodon milleri
Всего, учитывая синонимичные и спорные наименования, известно 20 видов диметродонов из 50 предполагаемых. Многочисленные останки из Техаса и Оклахомы до сих пор не описаны и не определены к какому-то определённому виду.
- D. teutonis Reisz & Berman, 2001. Нижние слои верхнего красного лежня (вольфкамп), Германия, Бромакер и Россия. Самый маленький диметродон, весил около 24 кг при длине в 0,8 метра. Единственный достоверно известный вид за пределами Северной Америки. Обладал высоким парусом.[4] Был привязан к сухопутной биоте.
- D. milleri Romer 1937. Сакмарский ярус, длина до 174 см, формация Putnam, Техас. Известен по двум скелета: почти полному MCZ 1365 и более крупному, но не так хорошо сохранившемуся MCZ 1367. Самый ранний вид диметродона из Техаса. В длину вырастал до 1,8 метров. Отличается от остальных видов строением невральных шипов: у D. milleri они имеют округлую форму, в то время как у остальных видов они напоминают восьмёрку в поперечнике. У этого вида позвонки также короче, чем у остальных. Череп высокий, морда короткая. Похожее строение имеют также D. booneorum, D. limbatus и D. grandis, возможно, что D. milleri был их предком. Близок к D. occidentalis.
- D. natalis Cope 1877. Сакмарский ярус, самый мелкий американский вид. Изображается с низким трапецевидным парусом, однако настоящая форма неизвестна. Длина черепа около 14 см, вес до 37 кг. Техас. Череп низкий, на верхней челюсти находились клыкообразные зазубренные зубы. Обнаружен недалеко от тела крупного D. limbatus. Син.: Clepsydrops natalis Cope, 1887.
- D. limbatus Cope 1877. Сакмарский и Артинский ярус — длина черепа до 40 см, общая длина до 2,6 м, из формации Admiral и Bell Plains в Техасе. Самый первый известный синапсид с парусом. Часто изображается в литературе. Первоначально был описан как Clepsydrops limbatus, к диметродону его отнесли Ромер и Прайс в 1940 году. Син.:Clepsydrops limbatus Cope, 1877; ? Dimetrodon incisivus Cope, 1878; Dimetrodon rectiformis Cope, 1878; Dimetrodon semiradicatus Cope, 1881.


Скелет D. incisivus
- D. incisivus Cope, 1878 — один из первых видов, иногда считается типовым. Возможный синоним D. limbatus.
- D. booneorum Romer 1937 — Артинский ярус — длина до 2,2 метра, Техас. Описан Ромером в 1937 году.
- D. gigashomogenes Case 1907. Кунгурский ярус. Достигал в высоту до 3,3 метров. Череп короткий и относительно высокий. Один из предков D. angelensis. Обнаружен в формации Арройо. Назван Кейсом ещё в 1907 году, до сих пор валиден.
- D. grandis Case, 1907. Ранний Кунгурский ярус. Достигал в длину до 3,2 м. Череп низкий, длиной 50 см. Имел всего четыре предклыковых зуба. Обнаружен в формации формации Арройо, Техас. Син.: Theropleura grandis Case, 1907; Bathyglyptus theodori Case, 1911; ? Dimetrodon gigas Cope, 1878; Dimetrodon maximus Romer, 1936; ? Dimetrodon cf. gigas grandis Sternberg, 1942.


Скелет D. grandis Case, 1907
- D. loomisi Romer 1937 . Кунгурский ярус. Вырастал до 2,5 метров. Обнаружен в формации Арройо, Техас. Отличается довольно низким черепом и зазубренной формой паруса.
- D. angelensis Olson 1962. Ранняя уфимская эпоха (поздняя Кунгурская). Последний и самый крупный известный вид. При жизни вырастал до 4—4,5 метров. Обнаружен в формации Сан-Анжело в Техасе. Череп длинный, до 50 см, и низкий, верхние клыки длинные тонкие. Все образцы плохо сохранились. Син.: ? Eosyodon hudsoni Olson, 1962 (nomen dubium), ? Steppesaurus gurleyi Olson & Beerbower, 1953.
- D. borealis Leidy, 1854. 270 млн лет назад, Остров Принца Эдуарда. Изначально был описан Д. Лейди по образцу верхней челюсти как самостоятельный род динозавров - батигнат (Bathygnathus borealis, в переводе "Тяжёлая северная челюсть"). Предполагалось, что высота челюсти превышала её длину. В голотипе сохранилось 7 крупных зубов разных размеров, высота фрагмента больше 10 см, длина — более 18 см. Задняя часть образца оказалась очень тонкой, так как при жизни она контактировала с другими костями. Челюсть покрыта сетью мелких бороздок. Зубы уплощённые, похожи на зубы варанов. На основе окаменелостей Лейди предположил, что песчнанники острова Принца Эдуарда датированы триасом, с чем не согласились многие геологи. Однако наличие "динозавра" говорило в пользу Лейди, пока в 1905 году Э. Кейз не доказал, что окаменелости батигната принадлежат сфенакодонту. Это позволило пределить возраст пород как раннепермский, о чём говорили обнаруженные остатки растений. Позднее была проведена реконструкция черепа батигната, которая оценила его длину в 40-45 см. В начале XXI века батигнат был синонимизирован с диметродоном.
- D. occidentalis Berman 1977 — единственный диметродон из формации Або/Катлер Нью-Мексико. Прелполагается, что достигал в длину 1,5 метра. Название означает «Западный диметродон». Известен по одному небольшому скелету. Предположительно связан с D. milleri.
- D. gigas Cope, 1878. Артинский и Кунгурский ярусы перми. Первоначально был описан как Clepsydrops gigas, однако позже его реклассифицировали как диметродона. К виду относится несколько хорошо сохранившихся образцов. Также рассматривается как синоним D. grandis.

")
")
")
")
")
")
")
")
")
")
")
- D. dollovianus Cope, 1888. Вид из артинского яруса Техаса. Известен по позвонкам и частям посткраниального скелета вместе с неполным черепом. Син.: Embolophorus dollovianus Cope, 1878; Dimetrodon fritillus (Cope, 1878).
- D. macrospondylus Cope, 1884. Вид из артинского яруса Техаса и Оклахомы, известен по позвонкам. Длина оценивается в 1,5 метра. Син.: Clepsydrops macrospondylus Cope, 1884; Dimetrodon platycentrus Case, 1907.
- D. kempae Romer 1937. Кунгурский ярус. Описан Ромером в 1937 году на основе плечевой кости и нескольких позвонков. Ромер и Прайс в 1940 году предположили, что D. kempae может являться самостоятельным родом.
Виды, выделенные в самостоятельные рода


Секодонтозавр, ранее считавшийся D. longiramus
- D. cruciger — описан Копом в статье 1878 году. Позже выделен им в род наозавр. Вид отличался от остальных диметродонов небольшими шипами на нервных шипах позвоночника. Позже наозавра синонимизировали с эдафозавром.
- D. longiramus — описан Кейсом в 1907 году. В 1940 году Ромер и Прайс заметили сходство D. longiramus с другим образцом, который также показывал удлинённую челюсть. Позднее этот вид выделили в род секодонтозавр.
Систематика


Диметродон является одним из ранних представителей класса синапсиды, дальних предков млекопитающих. Очень часто диметродона причисляют к динозаврам, а синапсид, особенно в старых источниках, называют зверообразными рептилиями. Оба эти утверждения неверны: диметродон обитал за 40 млн лат до первых динозавров, а синапсиды являются сестринским таксоном по отношению к рептилиям, отличной боковой ветвью. Диметродон, как и любой другой сфенакодонтид, стоит ближе к млекопитающим, нежели любой динозавр.
Согласно систематике, последний общий предок завропсид был тесно связан с пеликозаврами, поэтому при создании клад, включающих в себя и завропсидную, и синапсидную ветви, необходимо учитывать пеликозавров, у представителей которых присутствуют признаки и синапсидной ветви (сфенакоморфы и офиакодонты), и завропсидной (казеазавры и варанопиды).
В рамках синапсид диметродон входит в подотряд сфенакодонтов, выделенный вместе с эдафозаврами (с которыми образуют кладу сфенакоморфов) и офиакодонтами Ромером и Прайсом в 1940 году. Все три группы происходят из позднего карбона-ранней перми Европы и Америки. Отличить их можно по строению посткраниального скелета. Офиакодонты напоминали больше ящериц, из-за своей примитивности, и вели полуводный образ жизни. Эдафозавриды были более наземными животными, с полукруглыми парусами. Сфенакодонты были самой развитой из всех групп, вели наземный образ жизни, а особенности скелета сближали их с терапсидами — предками млекопитающих. Ромер и Прайс также выделили группу варанопсеид — более примитивных сфенакодонтид. Авторы посчитали их близкими родственниками, хотя варанопсеиды были меньше других сфенакодонтов и не имели парусов.


Современная классификация синапсид была предложена Робертом Рейсзем в 1986 году. Он основал новую классификацию на особенностях черепа, а не посткраниального скелета. Теперь варанопсеиды стали более примитивными синапсидами, некоторые учёные даже сближают их рептилиями. Диметродон остался членом сфенакодонтов в составе семейства сфенакодонтиды, одним из близких родственников терапсид. Кроме диметродона к этой группе относится его близкий родственник сфенакодон, обладающий удивительными челюстями секодонтозавр, крупный синапсид ктенорахис, более мелкие неозавр, ктеноспондил и катлерия, каменноугольный криптовенатор и спорные таксоны — батигнат и степпезавр, которых иногда синонимизируют с диметродоном. К сфенакодонтидам изредка относят и тетрацератопса, однако систематика этого рода очень спорна.
Ниже дана кладограмма филогенетического анализа Бенсона 2012 года:
| Amniota |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Однако анализ Бринка 2015 года даёт другие результаты:
| Sphenacodontidae |
| ||||||||||||||||||||||||||||||||||||
Синонимы


- Embolophorus Cope, 1878
- Theropleura Cope, 1880
- Bathyglyptus(?) Case, 1911
- Eosyodon Olson, 1962
Видовые синонимы
- Clepsydrops limbatus Cope, 1877
- Clepsydrops gigas Cope, 1878
- Clepsydrops natalis Cope, 1878
- Dimetrodon rectiformis (Cope, 1878) — описан на основе нескольких нервных шипов. Кейс переназначил этот вид в качестве синонима D. limbatus.
- Embolophorus dollovianus Cope, 1878
- Dimetrodon semiradicatus (Cope, 1881) — синонимизирован Кейсом с D. incisivus.
- Clepsydrops macrospondylus Cope 1884 — синоним D. macrospondylus.
- Dimetrodon platycentrus (Case, 1907) — синоним D. macrospondylus.
- Theropleura grandis Case, 1907 — синоним D. grandis.
- Bathyglyptus theodori Case, 1911 — синоним D. borealis.
- Dimetrodon maximus Romer, 1936
- Eosyodon hudsoni Olson, 1962
Палеобиология
Назначение паруса
С самого открытия паруса палеонтологи мучаются функцией паруса. Выдвигаются различные теории, однако ни одна из них не имеет точного подтверждения. Некоторые учёные считают, что парус мог служить средством маскировки или стабилизатором при ходьбе.
Терморегуляция
| Благодаря «парусу» времени на это занятие у диметродонов уходило сравнительно немного: если существо аналогичных размеров, но без «паруса», вынуждено было бы валяться на солнышке не меньше трёх-четырёх часов, чтобы повысить температуру тела хотя бы на несколько градусов, этому хищнику было довольно срока вполовину меньше, и многие другие обитатели поймы ещё только начинали переходить к дневной активности, а «живой парусник» уже спустился со своего древесного насеста и, опустившись в воду, неторопливо поплыл на соседний берег, изгибаясь всем своим длинным буро-полосатым телом. |


Возможно, парус мог служить средством терморегуляции, как у спинозавра. Эту теорию впервые предложили Ромер и Прайс в 1940 году. Подсчитано, что 200-килограммовый хладнокровный диметродон разогревался бы без паруса с 26 °C до 32 °C за 205 минут, а с парусом — за 80 минут. Таким образом, сфенакодонтид мог нагреваться и вести активную охоту уже раньше других хищников, или атаковать добычу до её пробуждения. Проходя через парус, кровь отводила избыток тепла, или наоборот нагревалась. В 1986 году Стивен Хак повысил сроки нагрева до 4 часов, основываясь на некоторых аспектах физиологии диметродона. Он также разработал модель, согласно которой парус мог эффективно нагревать тело синапсида утром и поддерживать температуру в течение дня, однако для выделения тепла он не годился и не позволял сохранять температуру в ночное время суток. Тем не менее, в 1999 году с использованием компьютерных технологий была разработана новая модель, показавшая, что парус удачно подходил и для сбора, и для выделения тепла в любое время суток и года.
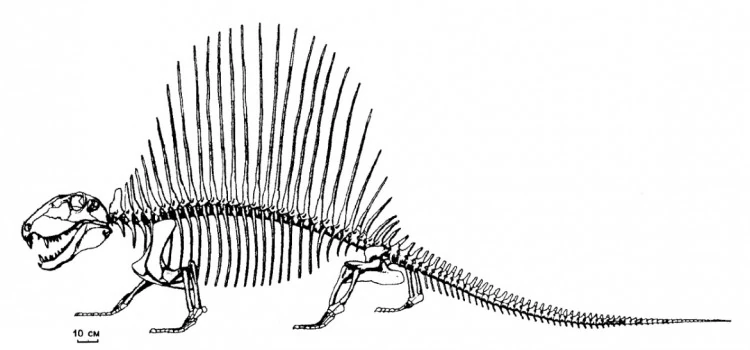
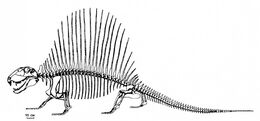
Прорисовка скелета D. limbatus
Большинство исследований предполагают, что парус был средством нагрева и охлаждения из-за того, что диметродон, как и другие ранние синапсиды, был либо хладнокровным, либо пойкилотермом, то есть зависел от температуры окружающей среды. А из-за массивного тела диметродон обладал большей тепловой инерцией, из-за чего изменения температуры у него происходило медленней, чем у мелких животных. Когда диметродон только просыпался, его добыча уже была активна, поэтому многие палеонтологи, в том числе и Хакк, предполагают, что парус помогал синапсиду быстро прогреться и вести активный образ жизни наравне с другими. А большая площадь паруса помогала быстрей охлаждаться.
В 1986 году Скотт Тернер и Ричард Трейси предложили, что парус диметродона мог быть свидетельством эволюции теплокровности у ранних предков млекопитающих. Согласно их теории, парус позволял сохранять хоть и низкую, но постоянную температуру тела. В отличие от млекопитающих, у которых температура регулируется повышенным метаболизмом, у синапсид, вроде диметродона и примитивных терапсид, теплокровность была гомеотермической, то есть, не слишком отличалась от такой же у рептилий. Например, современные гомеотермические ящерицы поддерживают температуру тела при помощи ферментов или поведения.
Недавние исследования подтвердили идеи Хакка о плохой теплоотдаче паруса и такому же поддержанию температуры. Кроме того, у близких родственников-сфенакодонтид, вроде сфенакодона или ктенорахиса, паруса вообще отсутствовали, на их месте были низкие гребни. А наличие паруса у мелких D. milleri и D. teutonis не соответствует теории о терморегуляции, так как в таком случае они бы отдавали больше тепла, чем получали, а некрупные размеры позволяли им самим удерживать необходимое количество теплоты. Некоторое время назад высказывались гипотезы, что паруса произошли от низких гребней или являлись наоборот, признаком примитивных видов.
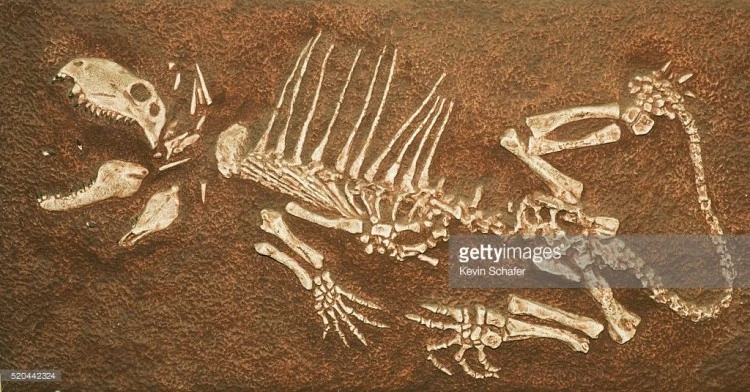

Скелет молодой особи диметродона
Интересно, что паруса соблюдают положительную аллометрию — у крупных диметродонов были крупные паруса. Это могло принести пользу при терморегуляции, так как площадь тела увеличивалась быстрее массы. Крупные животные вырабатывают больше тепла, поэтому им для охлаждения нужна соразмерная площадь. Однако исследования аллометрии 2010 года показали интересные особенности: оказалось, что парус рос быстрее, чем сам диметродон, из-за чего была предложена другая версия его назначения.
Привлечение внимания
У диметродона парус вполне мог играть роль органа привлечения внимания: различная форма, различная окраска могли использоваться во время брачных игр синапсид. Подобную конструкцию с такой же целью используют некоторые ящерицы, а также попозауроиды вроде ктенозавриска, аризоназавра и лотозавра и динозавры наподобие конкавенатора.
Половой диморфизм
| Образец AMNH 4636(внизу) и скелет из Мичиганского университета. Половые различия видны на наглядном примере |




У диметродона мог быть половой диморфизм. Некоторые останки относят к самцам, так как у них более толстые кости, более длинные черепа и увеличенные паруса. Такие черты наблюдаются у образца AMNH 4636, находящегося в Американском музее Естественной истории. В то же время, образец MCZ 1347 и несколько скелетов, расположенных в Музее Естественной истории Мичиганского университета, являются женщинами.
Слух[5]


Отличительной чертой сфенакодонтов является отражённая пластинка на углу нижней челюсти. Эта кость предназначалась для восприятия звуков. У большинства ранних тетраподов слуховое восприятие было плохо развито, а важнейшая из слуховых костей — стремечко — входила в состав основания черепа. Звук они воспринимали, припадаю головой или всем туловищем к земле. Сфенакодонты же были хищниками и при охоте держались голову высоко над землёй. Припадать к почве они не могли, так как конечности у них были длинными, и для того, чтобы использовать метод своих предков, сфенакодонты должны были ставить себя в невыгодные условия при приследовании добычи. Поэтому им пришлось формировать новый метод восприятия звуков, результатом которого стало появление среднего уха.
Ихнология[6]
| Когда вы видите ископаемый отпечаток, это живое ископаемое, и оно показывает, что это животное действительно ходило здесь, вот как оно ступало, вот где оно ходило. |


Лаура со своей находкой


Отпечаток ноги диметродона
В мае 2020 года на Острове принца Эдуарда геолог Лаура Макнил обнаружила дорожку следов, принадлежащую виду D. borealis. Первоначально она не верила, что нашла отпечатки лап, однако потом сообщила о находке доктору Джону Колдеру, геологу из Университета Святой Марии, чтобы задокументировать открытие. Колдер заявил, что находка принадлежит диметродону, позднее указав на важность данного открытия. Так как окаменелости находят в разных частях острова, находка отпечатков лап указывает, что Остров принца Эдуадра содержит одни из самых богатых свидетельств пермской фауны Канады.
По словам Лауры Макнил, находка будет выставлена в Национальном парке Острова принца Эдуарда летом 2020 года.
Палеоэкология
| То, что для одних становилось трагедией, другим обеспечивало безбедное существование, и в ближайшие пару месяцев на речных берегах будет не протолкнуться от хищников, так что даже взрослые диметродоны, обычно не жалующие соседей, вполне мирно уживутся на одной территории, не особенно порываясь отхватить у сородича половину хвоста! |


Диметродон известен из многих штатов — Техас, Оклахома, Юта, Нью-Мексико, Аризона, Огайо, а также Германии и Калининградской области. Тогда они все составляли континент Евроамерика. В приделах США окаменелости диметродона в основном находят в формациях Мирный ручей, Клир Форк и Вичито. В основном останки находят на месте болотистых местностей.
Наиболее часто останки диметродона находят в Красных грядах Техаса. Кроме него, очень часто встречаются также амфибии — архерия, сеймурия, диплокауль, эриопс и тримерорахис, рептилия капторин, а также синапсиды эдафозавр и офиакодон. Присутствовали крупные насекомые, служившие пищей для мелких животных. Эту характерную для юго-запада США фауну Олсон назвал «Пермокарбоновой хронофауной», доминировавшей в Северной Америке в раннюю пермь. Представители этой группы обитали в дельтах рек со свежей растительностью. В своей системе диметродон наверняка занимал позицию апексхищника.


Dimetrodon grandis от художника Франца Энтони
В окрестностях города Сеймур в штате Техасс была найдена линза, образовавшаяся на месте речной поймы. В ней было обнаружено огромное скопление пермских животных, среди которых было 39 диметродонов. Причём их предполагаемая добыча — эдафозавры и диадекты — была обнаружена всего в количестве нескольких штук. Чтобы поддержать такое количество апексхищников, этого было недостаточно. Однако удалось обнаружить останки 134 ксенакантид и 88 диплокаулюсов. Они значительно уступали диметродону в размерах, однако могли служить идеальным источником пищи.
Была предпринята попытка реконструировать процесс охоты диметродона. Передними крупными зубами он хватал добычу и вытаскивал её из воды резким движением головы и шеи с корпусом. На эту мысль наталкивают многочисленные прогрызенные черепа диплокаулей, однако мяса на них было мало, поэтому диметродоны поедали тела амфибий. Другая добыча — пресноводные акулы-ксенакантиды — могли оказывать сопротивление: найдены останки диметродонов со следами от зубов акул, однако обнаружено также 60 ксенакантов, перемешаннх с зубами диметродона. Обнаружен даже хрящ акулы с тремя зубами синапсида. Вполне возможно, что сфенакодонты охотились на рыб, обитавших в пересыхающих водоёмах.
Олсон предположил, что диметродон мог вести полуводный образ жизни, однако микроанатомия его костей указывает на чисто наземный образ жизни.


Найденный за пределами США D. teutonis обитал в совершенно иных условиях. Его останки были обнаружены в Тамбахской формации в районе Бромакер. Не удалось обнаружить много крупных хищник тетраподов, а размеры D. teutonis не позволяли ему охотиться на крупных диадектоморфных животных, в изобилие водившихся на этой территории. Всего на роль крупных хищников претендовало только три животных — собственно, D. teutonis, варанопсеид тамбакаринфекс и рептилиоморф сеймурия. Обнаружены также следы чуть более крупных сфенакодонтов, однако они могли быть оставлены и D. teutonis. Стоит учитывать, что на территории США диметродоны были равнинными жители, а на месте региона Бромакер в то время была гориста местность. В таких условиях крупные хищники не могли существовать, так как в основном они зависели от крупных амфибий. Зато для растительноядных животных здесь был рай — удалось обнаружить останки диадекта, казеида, оробата, следы арэосцелидовых или проколофоновых рептилий. Также нашли останки рептилий — тюринготириса и эвдибама, которые занимали ниши мелкого хищника и мелкого растительноядного соответственно. Они делили эти места с трематопидами ротарием и тамбахией. Кроме того, удалось обнаружить останки и следы жизнедеятельности медуз, жаброногих раков, червей, различных насекомых.[7]
В культуре


- В документальном фильме «Прогулки с монстрами» показаны диметродоны вида D. limbatus. Что странно, они показаны обитающими в Германии.
- В фильме «Путешествие к центру Земли» (1959) диметродонов изображали игуаны.
- Появляется в игре «Jurassic World: The Game» как один из легендарных хищников.
- Диметродон появляется в мультсериале «Земля до начала времён», что довольно нелогично — он вымер гораздо раньше любого из динозавров.


- В Перми был выпущен необычный чайный сервиз, на котором изображены пермские животные на фоне города. Так, камакопс изображён на фоне городской набережной за столиком, а диметродон в стиле XIX века прогуливается по городу. Цена такого сервиза — 400 рублей.[8]
- Кроме того, в некоторых поликлиниках пермской области на стенах можно увидеть диметродона и скутозавра, хотя они и обитали в разное время.[9]
- Диметродон появился в новом обновлении в игре «ARK: Survival Evolved» вместе с жуками-навозниками.
- Диметродон появляется в книге «В дебрях времени».
- Диметродон упоминается во множестве изданий, посвящённых пермской фауне.
Источники
- ↑ https://en.wikipedia.org/wiki/Dimetrodon
- ↑ https://age-of-mammals.ucoz.ru/index/dimetrodony/0-907
- ↑ http://www.skeletaldrawing.com/home/21stcenturydimetrodon
- ↑ https://ru.wikipedia.org/wiki/Диметродоны
- ↑ Звероящеры и другие пермские монстры. Д. В. Богданов. Издательство «Фитон XXI»
- ↑ https://www.cbc.ca/news/canada/prince-edward-island/pei-fossil-footprints-discovered-cavendish-park-1.5027811
- ↑ https://en.wikipedia.org/wiki/Tambach_Formation
- ↑ https://59.ru/text/business/66249604/
- ↑ https://59i.ru/novosti/novosti-permi-i-permskogo-kraja/polikliniki-permskogo-kraja-stanut-bolee-komfortnymi-i-poluchat-novyi-firmennyi-stil.html
Видео

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Галерея
- Основная статья: Диметродон/Галерея