![]()
Наносмил - небольшой кинжалозубый нимравид с зазубренными постклыковыми зубами из раннего олигоцена Северной Америки, размером приблизительно с рыжую рысь (Lynx rufus).
Известен только из местоположения CP 99A локации SX-I7, 11,5 миль к северу и 8,3/4 мили к западу от Кроуфорда, Sioux County. Формация Brule группы White River (штат Небраска, США).
Сравнительная анатомия[]
Череп узкий; затылок скошен назад, длина 14.6 мм. Почти горизонтальный с низким сагиттальным и затылочными гребнями; сокращенные назальные кости и очень антериорно размещенное постериорное палатальное отверстие. Лицевые кости менее расширены, чем у Hoplophoneus и Eusmilus, заканчиваются сзади чуть выше постериорного края C1; уровень гленоидной ямки на линии дёсен и вентральной границей мастоидного процесса. Сагиттальный гребень разделён V-образно выше гленоидной ямки (отклоняющийся антериорно гленоидного аспекта в Eusmilus и Hoplophoneus); вершина затылочного гребня на уровне с лицевыми костями (намного выше в Hoplophoneus); лицевые кости, не расширены настолько, как у Hoplophoneus или Eusmilus; постериорный край назальных костей с лобно-верхнечелюстными швами, как у Eusmilus; премаксиллярные кости, заканчиваются выше постериорного края клыков (сходство с Eusmilus в этом отношении), у Hoplophoneus же есть более удлиненное предверхнечелюстное завершение за постериорным краем клыка; орбиты большие; инфраорбитальный канал большой и покрыт костной тканью; резцовое отверстие простирается от фронта до задней части клыковых альвеол; углубление для постериорного палатального отверстия через постериорный край резцового отверстия, с дырой через P2.
Внутренние части ноздревых костей открываются антериорно вблизи M1; глубокая ямка, расположенная антериорно относительно M1 простирается вперед приблизительно до P4; антериорный край премаксиляррных костей отсутствует наряду с резцами и альвеолами резца; клык длинный и тонкий; однокоренной; P1 большой и двукоренной, маленький протокон, паракон отклонен назад, паракон и метакон заметно разделены; хищнический зуб с заметным парастилем, протокон уменьшенный, но поддерживается корнем, хищнический зуб сношен в срединном аспекте; M1 уменьшенный; орбитальные отверстия и швы отделены (у Eusmilus открыты сообща); круглое отверстие маленькое и расположено постериорно у вентрального края орбитального шва; имеется алисфеноидный канал; овальное отверстие открыто антеровентрально; большое отверстие постгленоидно; наличествуют мыщелковое отверстие и каротидное отверстие ("нижняя петрозальная пазуха").
Зубная формула ; редуцированный протокон и маленький парастиль на P4; P3, двукоренной с маленьким антериорным бугорком; p4 с большим параконидом; ml с маленьким метаконидом; фланец на нижней челюсти развит слабо. На М1 талонид почти отсутствует.

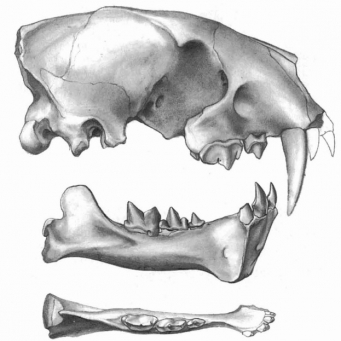
{kind=link}
Нижнечелюстная ветвь с очень маленьким фланцем, с единственным небольшим антериорным отверстием; единственное ментальное отверстие на боковой стороне фланца; диастемальный тонкий гребень; глубокая массетерная ямка ориентирована под М1; короноид короткий и округленный; угловой процесс уменьшенный с центральным вращением; I1 очень уменьшенный; I2 - загнутый постериорно; нижний клык подобен передним зубам; P1, P2 не обнаружены; P3 двукоренной с двумя выступами; P4 накладывается на антеро-латеральный край выступов М1, бугорки сжаты; carnassial с узкой меткой carnassial, протоконид М1 выше, чем параконид, метаконид М1очень маленький; M2 отсутствует.
Размеры черепа и челюсти UNSM 25505; антериорно-постериорная длина (AP), ширина (W); все размеры - в миллиметрах.
Размеры черепа:[]
- длина от постериорного конца клыковой альвеолы до постериорного поля мыщелка - 108.5;
- длина от постериорного конца гленоидной впадины до постериорного конца мыщелка - 35.6;
- минимальная ширина между высшими границами орбит - 26.8;
- ширина через посторбитальные сужения - 33.8;
- минимальная ширина посторбитальных сужений - 19.2;
- минимальная антериорная палатальная ширина между концами клыков - 17.3;
- ширина между постериорными концами альвеол для хищнических зубов - 43.1;
- максимальная ширина через парамастоидные кости - 44;
- максимальная ширина через затылочный мыщелок - 28;
- высота черепа от вершины затылочного пика до дорсальной границы большого затылочного отверстия - 32.1;
- ширина большого затылочного отверстия - 13.7;
- высота большого затылочного отверстия - 11.5;
- длина от антериорного конца клыковой альвеолы до постериорного конца альвеолы P4 - 41.8;
- длина от антериорного конца альвеолы P1 до постериорного конца P4 - 26.8;
- клыковые альвеолы AP - 10; W - 4.6;
- P3 AP - 9.4; W - 3.9;
- P4 AP - 16.4; W, 7.9;
- M1 AP - 4.3; W, 10.2;
- Нижнечелюстная ветвь: итог 85.8;
- расстояние от клыковой альвеолы собачьей до вентральной границы фланца - 22.3;
- минимальная глубина ветви - 17.9;
- глубина постериорной границы M1 - 14.9;
- толщина ниже M4 - 9.2;
- высота от внутренней границы угла до мыщелка - 15.7;
- высота от низшей границы угла до короноидного процесса - 17.1;
- поперечная ширина мыщелка - 13.7;
Зубы:[]
- I1 AP - 2.7, W - 1.5;
- I2 AP - 4.5, W - 2.4;
- I3 AP - 5.1, W - 4.1;
- нижняя клыковая альвеола AP - 5.7; W - 3.1;
- P3 AP - 6.8, W - 3.1;
- P4 AP - 10.6, W - 5.1;
- M1 AP - 5.8, W - 14.6.
Филогенетическое положение[]

{kind=link}
Североамериканский Eusmilus ограничен концом олигоцена (поздний Whitneyan и Gcringian). Nanosmilus значительно старше (ореллан) и более примитивен. Сопоставление между этими родами показывает следующие изменения от Nanosmilus до Eusmilus: развитие вертикальности затылочной области: снижение гленоидной ямки; расширение парамастоида за счёт параокципитального процесса; потеря P2, редукция P3, потеря нижнего P3; понижение короноидного процесса и развитие большого фланца нижней челюсти. Всё это - типичные эволюционные изменения у саблезубых плотоядных (Мартин, 1984).
Ассоциация голотипа Nanosmilus с Eusmilus уже была признана Шульцем и др. (1970), где Eusmilus демонстрирует предварительную ассоциацию с Dinictis. Nanosmilus, во многих характерах его расположения зубов, черепа и нижней челюсти, находится на уровне Dinictis и это во многих отношениях более примитивно (слабое развитие фланца, наклонный затылок, хорошо развитые премоляры), чем Hoplophoneus. Хорошо развитые парамастоидные процессы ясно показывают его сходство с гоплофонеусом. Таким образом, Nanosmilus - до некоторой степени самый примитивный известный член этого подсемейства и линии эусмилин, должно быть, отделившийся очень рано, поскольку эволюционно более продвинутые разновидности Hoplophoneus известны с чадрония.
Образ жизни[]
Большая часть североамериканской фауны олигоценовых парнокопытных представлена вымершей группой селенодонтных ореодонтов. Кажется вероятным, что последние формировали существенную часть добычи всех нимравидных плотоядных животных Северной Америки и, должно быть, являлись специальной добычей миниатюрного Nanosmilus. Несмотря на небольшой размер, вполне допустимо, что Nanosmilus был небольшим свирепым хищником, который был в состоянии охотиться на животных размером с домашнюю свинью или оленя (Мартин, 1992).
Этимология[]
Nanos (греч.) «небольшая (карликовая) и smile – сабля (разделочный нож)». Видовое название – в честь Bjorn Kurten, учитывая его вклад в изучение эволюции плотоядных животных.